 Takk, professor Lenski, LTEE gjør det bra!
Takk, professor Lenski, LTEE gjør det bra!
(Long-Term Evolution Experiment (LTEE) -her)
Michael Behe; 29. mars 2019
Oversatt herfra. {Understrekinger og kursiveringer er ved oversetter.}
Bilde 1: Richard Lenski, av Zachary Blount [ CC BY-SA 4.0 ], fra Wikimedia Commons.
Dette er det tredje i en serie innlegg som svarer på den utvidede kritikken av 'Darwin Devolves' fra Richard Lenski på bloggen hans, Telliamed Revisited. Professor Lenski er kanskje den mest kvalifiserte forskeren i verden til å analysere argumentene fra boken min. Han er Hannah Distinguished Professor of Microbial Ecology ved Michigan State University, en MacArthur ('Genius Award') Fellow, og medlem av National Academy of Sciences med hundrevis av publikasjoner. Han har også en sterk interesse for vitenskaps historie og filosofi. Hans eget laboratorie-evolusjonsarbeid er et sentralt tema i boken. Jeg er veldig takknemlig til professor Lenski for å ta tid til å vurdere 'Darwin Devolves'. Hans kommentarer vil tillate interesserte lesere raskt å måle den relative styrken av argumenter mot bokens budskap.
 Uforutsette konsekvenser
Uforutsette konsekvenser
Jeg har allerede tatt opp flere av problemene Lenski har reist på sin blogg i sitt tredje innlegg om Darwin Devolves, 'Is LTEE Breaking Bad?' -her, I mine svar ( her, her og her) til gjennomgang av mine Lehigh-kolleger, fordi de ofte siterte fra hans arbeid. Likevel er repetisjon et fint læringsverktøy. Så her vil jeg igjen snakke om disse problemene og også ta opp noen andre.
I 'Is LTEE Breaking Bad?' er Lenski enig i at de gunstige mutasjonene som er sett i hans Long Term Evolution Experiment, overveiende er nedbrytende eller noen som innehar tap av funksjon. Likevel, bekymrer det ham ikke fordi "LTEE representerer et ideelt system for å observere nedbrytende evolusjon." Fordelaktige nedbrytende endringer er bare å forvente der, virker det som.
-LTEE ble designet (intelligent, etter min mening!) til å være ekstremt enkelt for å ta opp noen grunnleggende spørsmål om dynamikken og repeterbarheten ved evolusjon, samtidig som komplikasjoner minimeres. Det var ikke ment å etterligne naturens kompleksitet, og det var heller ikke ment å være et teststed for utvikling av nye funksjoner. Miljøet der bakteriene vokser er ekstremt enkelt. ...
-Faktisk er LTEE-miljøet så ekstremt enkelt at man med rimelighet kan forvente at bakteriene vil utvikle seg ved å bryte ned mange eksisterende funksjoner. Det er fordi cellene, uten konsekvens, kunne miste sine evne i å utnytte ressurser som ikke er til stede i kolbene, miste sitt forsvar mot fraværende angripere og konkurrenter, og miste sin evne til å motstå ikke-lenger-relevante ekstreme temperaturer, gallesalt -her, antibiotika og mer. [Uthevet i originalen.]
 Med andre ord er det mange verktøy i den robuste E. coli genomiske verktøykassen som ikke ville være nødvendige i Michigan State Lab. Det kan miste dem uten umiddelbar konsekvenser. Faktisk kan det til og med være en fordel å miste dem, enten ved bare å spare energi ved å lage dem, eller ved å omdirigere ressurser til andre stier som er mer brukt i laboratoriemiljøet.
Med andre ord er det mange verktøy i den robuste E. coli genomiske verktøykassen som ikke ville være nødvendige i Michigan State Lab. Det kan miste dem uten umiddelbar konsekvenser. Faktisk kan det til og med være en fordel å miste dem, enten ved bare å spare energi ved å lage dem, eller ved å omdirigere ressurser til andre stier som er mer brukt i laboratoriemiljøet.
Likevel er en uunngåelig konsekvens av å miste gener, å miste fleksibilitet - mens den overordnede bakteriestammen kunne utvikle seg til å leve i laboratoriemiljøet, kan ikke mutantbakteriene gå hjem igjen. Hvis deres miljø skiftet tilbake til det mer komplekse, mer fiendtlige som de hadde blitt tatt fra, ville de ikke lenger være konkurransedyktige. Som en sterk regel, forventer vi situasjoner der noen arter - ikke bare E. coli - enten i laboratoriet eller i naturen, tilpasser seg ved å miste gener samtidig som de begrenser sin evne til å tilpasse seg fremtidige miljøforandringer. Akkurat som lab E. coli blir mer miljømessig begrenset ettersom det mister flere gener, er også eksemplene fra naturen om isbjørnen og Yersinia-pesten, som jeg sitert i Darwin Devolves, det.
Super fart
Som nevnt ovenfor satte professor Lenski opp LTEE for å undersøke 'grunnleggende spørsmål om dynamikken ... ved evolusjonen.' En grunnleggende dynamikk som resultatene viser med stor klarhet er at fordelaktige nedbrytende mutasjoner forekommer størrelsesordener raskere enn gunstige ikke-nedbrytende. Fordelaktig nedbrytende mutasjoner som ødeleggelse av ribose-operonen skjedde på om lag hundre generasjoner -her. På den annen side oppstod den sene ikke-nedbrytende citrat-mutasjonen først etter 30 000 generasjoner. Når det gjelder ren vekst i laboratoriemiljøet, hjalp det bakteriene mye mer enn noen av de nedbrytende mutasjonene- citratmutantene tok helt over kolonien i løpet av natten. Ikke desto mindre virket citratmutasjonen bare godt etter at kanskje et dusin mutasjoner i det istykkerbrutte gener allerede hadde feid over populasjonen for fiksering, noe som permanent begrenset bakteriestammen.
Hvorfor er det slik? Citratmutasjonen var mye langsommere i å dukke opp, selvfølgelig, fordi det krevde mer krevende, mindre sannsynlige forhold som nødvendigvis opptrer mye sjeldnere. Som regel vil alle ikke-nedbrytende mutasjoner være mindre hyppige enn nedbrytende. Så et svært viktig dynamisk resultat som LTEE gjør krystallklart er at nedbrytende mutasjoner kommer veldig raskt, konstruktive kommer mye langsommere. Med andre ord, vil alle nyttige degraderende endringer ha tiden de trenger for å ta over en populasjon, godt før ankomsten av bestemte ikke-nedbrytende endringer, som for eksempel citratmutasjonen.
 Betydningen av Citrat
Betydningen av Citrat
Professor Lenski mener at citratmutasjonen er et godt eksempel på den konstruktive kraften ved evolusjon. Likevel betrakter han den isolert og overser den overveldende nedbrytende konteksten omkring den hendelsen.
-Til sin kreditt skriver Behe om linjen som utviklet evnen til å konsumere citratet. Men han avviser den som en "sideshow" (s. 365 [ sic, faktisk s. 190]), fordi han nekter å kalle denne nye evnen en gevinst ved funksjon. I stedet skriver Behe (s. 362 [ sic, s. 189]) at under sin selvoppfyllende ordning "vil mutasjonen bli regnet som modifikasjon av funksjonen - fordi intet nytt funksjonelt kodet element ble vunnet eller gikk tapt, bare kopiert." Med andre ord, vil Behe ikke telle noen nyutviklet funksjon som en funksjons-gevinst, med mindre et helt nytt gen eller kontrollregion 'poff' kommer til eksistens. [Utheving i originalen.]
Jeg mente ikke å såre Professor Lenskis følelser med 'sideshow' bemerkningen om citrat-mutasjonen. Jeg er mer enn glad for å være enig i at det var en veldig interessant utvikling i lab-utviklingsprosjektet. Imidlertid, som jeg har skrevet før -her, kalte jeg det for et 'sideshow' fordi det var mange forutgående nedbrytende mutasjoner, etterfulgt av flere som forbedret effektiviteten. Sammenlignet med nedbrytende mutasjoner, ble den (citrat mutasjonen) numerisk overvunnet. Etter min mening, er langt det viktigste poenget at nyttige nedbrytende mutasjoner virker så fort, i forhold til konstruktive at de vil 'sverme' etter enhver forandring i miljøet, og raskt tilpasse seg en organisme. Det er akkurat det vi bør forvente fra den første regelen av adaptiv evolusjon. Hvis en heldig konstruktiv mutasjon - noen ganger, etter hvert, dukker opp, er det også fint, men det retter ikke opp skadene som er forårsaket av 'gift-pille' mutasjonene.
 LTEE er 'poff-løs'
LTEE er 'poff-løs'
Når det gjelder professor Lenskis klage om at "Behe ikke teller noen nyutviklet funksjon som funksjons-forsterkelse, med mindre noe helt nytt gen eller kontrollregion 'poffer' fram" (uthevelse fjernet), tar han feil. I både min Quarterly Review of Biology-artikkel og Darwin Devolves regnet jeg sigdcelle mutasjonen (en enkelt aminosyreendring) av hemoglobin som gevinst for FCT (FKT står for "Funktsjonelt-Kodet-elemenT"), fordi det resulterte i en nytt protein-protein bindings-sted. Som jeg utfyllende forklarte begge steder, klassifiserte jeg mutasjoner i henhold til deres molekylære effekter - ikke deres fenotypiske effekter - alt etter om de resulterte i gevinst eller tap av slike funksjonelt kodede elementer -her, som :
--promotorer; forsterkere; isolatorer; Shine-Dalgarno-sekvenser; tRNA gener; miRNA gener; proteinkodende sekvenser; organelle mål- eller lokaliserings-signaler; intron/extron spleisningssteder; kodoner som spesifiserer bindingsstedet for et protein for et annet molekyl (slik som dets substrat, et annet protein eller en liten allosterisk regulator); kodoner som spesifiserer et behandlingssted for et protein (slik som et spaltnings-, myristoylerings- eller fosforyleringssted); polyadenyleringssignaler; og transkripsjons- og oversettelses-termineringssignaler.
For korrekt å kunne evaluere Darwins mekanisme må man observere hva det gjør på molekylært nivå når det gjelder kodede genetiske elementer. Mange omhyggelige studier dokumenterer at det i overveiende grad nedbryter dem. Ingen nye slike elementer ble sett i LTEE.
Over 90 prosent degraderende kvalifiserer som 'hovedsakelig'
 Professor Lenski forsøker å skjerme minst noen av de tjenlige mutasjonene til LTEE fra å bli merket som nedbrytende.
Professor Lenski forsøker å skjerme minst noen av de tjenlige mutasjonene til LTEE fra å bli merket som nedbrytende.
-I Darwin Devolves hevder Behe (s. 344 [ sic, faktisk s. 179]) at "det er svært sannsynlig at alle de identifiserte fordelaktige mutasjonene virket ved å forringe eller rett og slett bryte respektive forfedre-gener." Han inkluderer en fotnote som anerkjenner vårt arbeid som foreslår finjustering av noen proteinfunksjoner, men der skriver han (s. 609): "Nyere undersøkelser fra Lenskis laboratorium antyder at mutasjoner i en liten minoritet (10 av 57) fra utvalgte E. coli- gener kanskje ikke fullstendig bryter dem, men, som de sier, 'finjusterer' dem (sannsynligvis ved å forringe deres funksjoner). "Hvorfor hevder Behe at finjustering av gener oppstod 'sannsynligvis ved å forringe deres funksjoner'?
-Kanskje er det fordi denne påstanden støtter hans krav, men mer imøtekommende, mistenker jeg at den underliggende årsaken ligner de problematiske konsekvensene som fikk Behe i vanskeligheter når det gjelder isbjørnens gener. Det vil si at hvis man antar at en opprinnelig tilstand i et gen er perfekt, det ikke noe rom for forbedring i funksjonen, og de eneste mulige funksjonelle endringene er nedbrytende. ... Det ville være overraskende om noen proteiner ikke kunne finjusteres slik at deres aktiviteter ble forbedret under spesiell pH, temperatur, osmolaritet og andre forhold i LTEE.
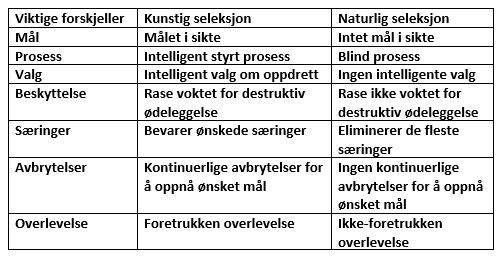 Flere kommentarer: Først, isbjørnen. Det viser seg at Lenski og hans upålitelige kilder grep til et halmstrå for å unngå den bentframme konklusjonen at nedbryting av isbjørnens APOB-protein faktisk hjalp den til å tolerere et høyt fettholdig kosthold -her. Den samme effekten er sett hos mus når en kopi av genet er slått ut.
Flere kommentarer: Først, isbjørnen. Det viser seg at Lenski og hans upålitelige kilder grep til et halmstrå for å unngå den bentframme konklusjonen at nedbryting av isbjørnens APOB-protein faktisk hjalp den til å tolerere et høyt fettholdig kosthold -her. Den samme effekten er sett hos mus når en kopi av genet er slått ut.
For det andre er jeg enig i at man bør forvente litt finjustering i LTEE. Likevel, hvis 'bare' 47 av 57 fordelaktige mutasjoner arbeidet med å nedbryte eller ødelegge gener i LTEE-prosjektet, ville jeg vurdere at argumentet mitt er helt rettferdiggjort. Tross alt hevdet jeg aldri at alle gunstige mutasjoner må være nedbrytende. Den første regelen for adaptiv evolusjon er probabilistisk, ikke preskriptiv.
For det tredje, grunnen til at jeg betraktet de gjenværende utvalgte proteinene til trolig å være nedbrytende, så kan årsakene finnes i Lenskis artikkel selv -her. Tre av de ti (nadR, pykF og yijC) som møtte teamets kriterium for å være potensielt finjusterende (med samme punktmutasjon valgt minst to ganger i forskjellige replikerte stammer) kunne også velges dersom de hadde tap-av-funksjon mutasjoner, som indikerer at nedbrytning av proteinets aktivitet var gunstig. Et annet protein (spoT) led av flere forskjellige mutasjoner, som sannsynligvis indikerer at det ble skadet på forskjellige punkter (som selvsagt er mye lettere å gjøre enn å forbedre et protein). Flere av de andre proteinene (atoC, hflB, infC, rpsD) kjøpte enkelt aminosyre-substitusjoner som, jeg er enig i, kan 'finjustere' proteinene i den forstand at de marginalt justeres til forholdene hos LTEE. Imidlertid illustrerer de også vanskeligheten ved å gjøre det, siden det tok titusenvis av generasjoner å dukke opp, mye lenger enn gunstige nedbrytende mutasjoner i andre gener.
Således, på slutten av tusenvis av generasjoner, er det noe håp om at kanskje 4 av 57 fordelaktige mutasjoner - færre enn 10 prosent - ikke bryter ned sine respektive gener. Som jeg skrev i Darwin Devolves, virker 'Darwins mekanisme hovedsakelig ved å kaste bort genetisk informasjon til fordel for kortsiktig gevinst.'
 Mauren, og gresshoppen
Mauren, og gresshoppen
En siste, illevarslende poeng. Artikkelen Lenski refererer til ovenfor, heter 'Core Genes Evolve Rapidly In The Long-Term Evolution Experiment with Escherichia coli.' Deler av Abstraktet lyder:
-Vi spurte om de samme genene som utvikler seg raskt i det langsiktige utviklings-eksperimentet (LTEE) med Escherichia coli, også har utviklet seg i stor grad i naturen. For å gjøre denne sammenligningen identifiserte vi ~ 2000 kjerne-gener delt i 60 E. coli- stammer. Under LTEE akkumulerte kjerne-gener betydelig mer ikke-likeartede mutasjoner enn fleksible (dvs. ikke-kjerne) gener. Videre er kjernegener under positiv seleksjon i LTEE mer konserverte i naturen enn det gjennomsnittlige kjernegenet.
Med andre ord vil tilfeldig mutasjoner like tankeløst kaste bort selv kjerne-gener (antagelig mer nyttig under flere omstendigheter) enn ikke-kjerne gener, dersom det gir en øyeblikkelig fordel. Det er ingen plan for, eller forutseenhet for, fremtiden i darwinismen. Som i fabelen om gresshoppen og mauren, er Darwins mekanisme tydelig gresshoppen.
I takknemlig anerkjennelse
Enhver som er interessert i emnet evolusjon, burde føle seg takknemlig for Richard Lenski og hans laboratorium. På grunn av deres arbeid kan vi diskutere implikasjonene av ekte, relevante fakta, snarere enn fra grunnløse spekulasjoner.
Oversettelse og bilder ved Asbjørn E. Lund